

The Japanese Dog: Branding and Mental Images
The concept of “the Japanese dog” is intriguing. Dogs are so much more than just dogs, and clearly this has...
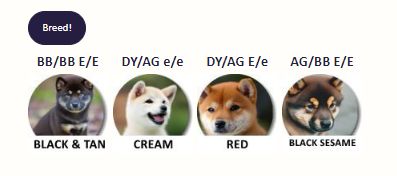
Shiba Inu Puppy Generator (International Version)
Here’s an English version of the coat color generator. This generator produces litters according to the current (2024) understanding of...
The First Shibas in Europe
The first known Shibas of Europe arrived in the early 1970s to the Swedish Kennel Manlöten owned by Inga and...
Shibas in gene test: colour loci
Written originally in Finnish for Shibasanomat 2/2020. The English version kindly translated by Topi Kuusinen. Update: MyDogDNA breed database as...
Viimeisimmät kommentit