

Mistä valkoinen väri on tullut nykyshiboihin?
Shiban valkoisen värin saa aikaan E-geenin mutanttiversio, jonka nimi on yksinkertaisesti e eli ”pikku-e”. Se periytyy peittyvästi verrattuna ”isoon E:hen”,...

Kommentit shikokujen kuuluisimmista kantakoirista
Shikokunkoiran sukupuun juuristoon kaivautuminen odottaa vielä aikaansa. NIPPO:n shikokusivun ja Shigeru Katon Nihonken-blogin perusteella sieltä löytyy tällaisia uroksia, jotka on...

Shikokunkoira ja geneettisen monimuotoisuuden yllätykset
Shikokunkoirille on tehty sama DNA:n monimuotoisuuskartoitus kuin shiboille. Itse asiassa niitä on tutkittu paljon enemmän kuin shiboja eli tällä hetkellä...

Shikokunkoirien kystinuria – totta vai tarua?
Blogin loppuvuosi on omistettu shikokunkoirille. NIPPO on nyt vedonnut länsimaiseen yleisöön siinä, että keskikokoiset japanilaisrodut saataisiin pelastettua sukupuutolta. Shikokuja rekisteröitiin...
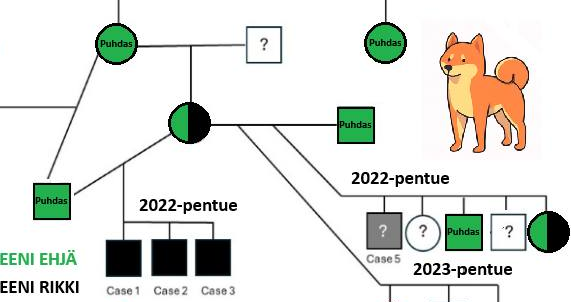
Uusi geenivirhelöytö shiboista
Shiboille on juuri todennettu kolmas rotukohtainen tautimutaatio. Kyseinen mutaatio DMD-geenissä johtaa vialliseen dystrofiini-proteiiniin. Seurauksena on lihasdystrofia. Sen karmein oire voi...

Japanilaisrodut mutaatiotestissä
Onko japanilaisrotuisen koiran laajasta geenitestauksesta hyötyä? Löysin kiinnostavan tutkimuksen, kun aloin kiinnittää huomiota ulkomaisten shibakennelien sivuilla toistuvaan DM-geenitestitulokseen. Mikä edes...

Ajatuksia Jalostuksen peruskurssilta
Kennelliitto järjesti kaksipäiväisen etäluentosarjan liittyen koirien jalostukseen. Minikurssi oli avoin kaikille. Sinne siis! Mulla on T-O-D-E-L-L-A kaksijakoinen suhtautuminen koko jalostusasiaan....

Akita ja söpön pörröisyyden hintalappu
Otsikko ei viittaa rahaan. Uusi nihonkentutkimus Japanista käsittelee akitan pitkäkarvamutaatiota ja sen yleistymistä kantajien takia. Koirilla pitkäkarvaisuus on jäljitetty FGF5-geenin...

Shiban dyskinesiageenin jäljillä?
Dyskinesiajutut on taas ajankohtaisia, ja paperilla aivan erisukuisen shiban takia. Samat geenit ja geenivirheethän niillä on ympäri maailman. Siitä inspiroituneena...

Shiban loppu vai uuden alku?
Elämmekö rotukoirien kanssa sittenkään niin tylsää vaihetta kuin ensinäkemältä vaikuttaa? Tykkään zoomailla ajassa ja paikassa, koska oon huomannut, että se...
Viimeisimmät kommentit